
Vastarel
Cheap vastarel 20mg without a prescription
It would not be sensible to delegate to each of our distracted free wills something that would be fatal if forgotten medicine mound texas discount vastarel 20mg amex. But, to a certain point, we can control our breathing consciously, its rhythm, volume, flow. Establishing this bridge between the implicit and the declarative turns out to be, as we will see, a key variable in every form of learning. People engaged in learning to type, for example, begin by searching out each letter with their eyes, exerting great effort and concentration. When we touch-type, our brains are somewhere else, reflecting on the text, talking to someone else or daydreaming. In other words, the learning curve grows until it reaches a value where it stabilizes. But, of course, this value is not the same for everyone; the world record is held by Stella Pajunas, who managed to type at the extraordinary pace of 216 words per minute. Yet by doing methodical, sustained exercises to increase our speed, all of us can improve substantially. The history of human virtue What happens in the example of typing speed occurs with almost everything we learn in life. After years of intense effort at school, many of us achieve reading quickly and with little effort. Yet if any of us revisited a methodical, sustained process, and devoted time and effort, we could significantly increase our speed without losing comprehension along the way. The narrative of learning in each of our life cycles is replicated in the history of culture and sport. In the early twentieth century, the fastest runners of the times achieved the extraordinary feat of running a marathon in two and a half hours. Some compositions by Tchaikovsky were technically so difficult that in his day they were never played. Today they are still considered challenging but there are many violinists who can play them. Violinists today manage to play those pieces because they can devote more hours to their practice, because the point at which they feel the goal is accomplished has changed, and because they have better training procedures. This is good news; it means that we can build on these examples to attain goals that today are inconceivable. Fighting spirit, on the other hand, feels more human because it is associated with will and the feeling that we could all achieve it. It is, in fact, an element of temperament, which refers to a vast term that defines personality traits including emotiveness and sensitivity, sociability, persistence and focus. As in a film by Richard Linklater, they meticulously followed the development of hundreds of children from different families, from the day they were born through adulthood. They measured nine traits of their temperaments: (1) Their activity levels and types. They found that while these traits were not immutable, they at least persisted to a striking extent throughout their development. Over the last fifty years, this foundational study by Chess and Thomas has been continued with a multitude of variations. If genes explain more or less half of our temperament, the other half is explained by the environment and social petri dish in which we develop. Of almost all cognitive variables, the most decisive factor is the home a child grows up in. Siblings are similar not only because they carry similar genes but also because they develop on the same playing field. Different studies on adoptions and twins show that the home contributes very little to the development of some aspects of temperament. Searching for the nature of human altruism, an Austrian behavioural economist, Ernst Fehr, has shown this quite conclusively for one of the foundational traits of temperament: the predisposition to sharing. When the younger siblings get something, they keep it for themselves in their jungle filled with older predators. All parents with more than one child recognize that the anxiety, fragility and above all ignorance in which a first child is raised are not repeated. As a result of this, some social aspects of young children, such as their disposition to share, do not depend so much on the home experience but are rather learned on other playing fields of life. This explanation is mostly based on the work of Stella Chess and Alexander Thomas, who have meticulously observed the persistence and malleability of different personality traits that make us what we are. But we will see later in this chapter that it is closely related to aspects of the motivation and reward system that define temperament and are a gateway to learning. What we perceive as talent is not an innate gift but rather, almost always, the fruit born of hard work. Somebody with this aptitude is usually considered a musical genius, so out of the ordinary as to be viewed as some sort of mutant, like an X-Man of music, gifted with a genetic package that gives them this unusual virtue. In fact, most children have almost perfect pitch, but without practice it atrophies. And children who begin training at a conservatory at a young age have a very high incidence of perfect pitch.
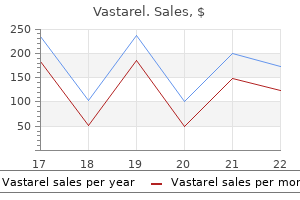
Best 20mg vastarel
Moreover treatment joint pain generic 20mg vastarel free shipping, the same cell continued to fre dur ing the second delay period if the response was directed to a specifc location. These results indicate that, in terms of stimulus atributes, cells in the prefontal cortex exhibit task specifc selectivit. Her knowledge of the minimal right afer the stimulus is presented and entirely Golden Gate Bridge requires activation of a distributed network absent during the delay period. Moreover, the response of cortical regions that underlie the representation of long-term of these cells is malleable. These cellular responses by themselves do not tell us successively for 1 s each during an encoding interval. The be that long-term representations are stored in the pre participants were instructed to remember only the faces. Patients ing the encoding interval, the processing demands on work with fontal lobe lesions do not have defcits in long ing memory were manipulated. This later hypothesis the onset of the encoding period, and this response was jibes nicely with the fact that the prefontal cortex is ex maintained across the delay period even though the scren tensively connected with postsensory regions of the tem was blank (Figure 12. The sus representation can be sustained through the interactions tained response during the delay period was greater when bet n prefontal cortex and posterior brain regions, the participant had to remember thre or four intact faces one that can facilitate goal-oriented behavior. This alternative hypothesis has ben examined in many By using faces, the experimenters could also compare functional imaging studies. In one representative study, activation in the prefontal cortex with that observed in researchers used a variant of a delayed-response task the fusiform face area, the inferior temporal (also called (Figure 12. After a delay period, a probe stimulus is presented and the participant indicates if that face was part of the memory set. The magnitude of this effect is related to the number of faces that must be maintained in working memory. When the stimuli were memory retrieval the peak response is slightly earlier presented, either during the encoding phase or for the in the prefontal cortex. During the delay sistent with the general tenets of the model sketched in period, as noted already, the prefontal response remains Figure 12. Following a de Spatial lay period, a series of pictures containing letters backward D and squares at various locations is presented. Dorsolateral prefrontal cortex is more Verbal G K active when the stimuli must be remembered in backward R reverse order, independent of whether the set is composed of locations or letters. When the judgment required that the items be remembered in the forward stream of stimuli. Participants are instructed to push a direction, study participants had to internally maintain buton when they detect a repeated stimulus. In the sim a representation of the stimuli using a natural scheme plest version (n 5 1), responses are made when the same in which the items could be rehearsed in the order pre stimulus is presented on to successive trials. The backward conditions were more challeng complicated versions, n can equal 2 or more. With n-back ing, because the representations of the items had to be tasks, it is not sufcient simply to maintain a representa remembered and manipulated. Activation A similar patern is observed in many imaging in the lateral prefontal cortex increases as n-back task studies, indicating that dorsolateral prefontal cortex is difcult is increased, a response consistent with the idea critical when the contents of working memory must be that this region is critical for the manipulation operation. One favorite testing variant for studying The n-back tasks capture an essential aspect of pre manipulations in working memory is the n-back task fontal function, emphasizing the active part of working (Figure 12. The recency task that patients with fontal lobe Goal-Oriented Behavior | 519 1-back condition 2-back condition a crude hierarchy: For the simplest of working memory tasks, the activation may be limited to more posterior K K prefontal regions or even secondary motor areas. When such contingencies are made even more challenging by changing the rules Response L Response T fom one block of trials to the next, activation extends A A even farther in the anterior direction (Figure 12. Tese P P complex experiments demonstrate how goal-oriented be havior can require the integration of multiple pieces of information. In the extreme, we might think Prefrontal Cortex of the most posterior part of the fontal lobe, the primary motor cortex, as the point where abstract The maintain-manipulate distinction ofers a processing intentions are translated into concrete movement. The fontal pole was the one region that was recruited upon presentation of the instruction cue in all four conditions (shown in Figure 12. It has ben proposed that the fontal pole is essential for integrating the specifc contents of mental activit into Posterior a general famework. For example, a response to a that they could volunter to work in the lab and run their letter might be made if the color of the letter was green, but not if own cool experiments in the future. For example, suppose it is the hotest day of summer Decision Making and you are at the lake. Our brains start making decisions as associate with your previous encounters with root ber soon as our eyes futer open in the morning: Do I get up foats. More posterior regions become active as you begin now, or stay cozy and warm for another hour Do root ber foat, and that goal becomes the center of work I skip class to take of for a wekend ski trip You also draw on dorsal regions that will be essential for Rational observers, such as economists and mathema forming a plan of action to drive to the A&W. For instance, the abilit to establish the sequence of actions required to why would someone who is concerned about eating accomplish your goal. The feld of neuroeconomics has emerged as an Goal-oriented behavior requires the retrieval, selection, interdisciplinary enterprise with the goal of explain and manipulation of task-relevant information. Many of their ideas can be tested both Working memory consists of transient representa with behavioral studies and, as in all of cognitive neuro tions of task-relevant information. The prefrontal cortex (especially the lateral prefrontal cortex) is a key compo science, with data fom cellular activit, neuroimaging, nent in a working memory network. This work also helps us understand the Physiological studies in primates show that cells in the functional organization of the brain. Normative decision theories picture is observed in functional imaging studies with defne how people ought to make decisions that yield the humans. Very ofen, however, such theories fail to Decision Making | 521 predict what people actually choose. Afer we repeat that action, and if the suboptimal choices present less of a mystery to evolution outcome is consistent, the process becomes habitual; that ary psychologists. Another dis evolution to optimize reproduction and survival in a world tinction can be made beten decisions that are model that difered quite a bit fom the one we currently occupy.
Buy vastarel 20mg fast delivery
These early songs may have faltering pitch medicine for high blood pressure buy 20 mg vastarel mastercard, irregular tempo, and notes that are out of order or poorly reproduced. However, sonograms of songs recorded over several weeks or months reveal that during this practice period the bird ne-tunes his efforts until he produces an accu 67 the Neural Basis of Song Learning in Birds rate copy of the memorized template. This process requires hearing oneself sing; birds are unable to reproduce memorized songs if they are deafened after memorization but before the practice period (Konishi 1965). Sensitive Periods for Song Learning In many species, referred to as closed-ended learners, song memoriza tion occurs during a restricted period of development, often within the rst few months after hatching. This sensitive period has been demon strated in the laboratory by presenting young birds with a series of taperecorded songs drawn from the dialects of their own species. In other species, referred to as open-ended learners, song learning reoccurs each year. For example, the repertoires of starlings (Sturnus vulgaris; Eens, Pinxten, and Ver heyen 1992) and canaries (Serinus canaria; Nottebohm and Nottebohm 1978) increase or change from year to year. Variation across species with respect to when songs are learned raises several interesting questions: what determines the length of the sensitive period for song learning The length of the sensitive period appears to be regulated by both external and internal factors. Young birds deprived of an opportunity to hear song will memorize at a later age than those that were tutored as edglings, indicating delayed closure of the sensitive period (Kroodsma and Pickert 1980; Slater, Jones, and TenCate 1993). Birds prevented from hearing the songs of their own species throughout this period will produce abnormal songs, as shown in gure 5. Once the bird begins to practice an abnormal song, subsequent tutoring will not improve it, indicating that irreversible changes have occurred in the brain. The Neurobiology of Sensitive Periods If a bird is able to learn to sing at one age but not another, we must con clude that some change occurs in the brain between these two time 68 Carol Whaling points. It should be possible, if we knew where to look, to nd neural changes that correspond to the duration of the sensitive period. Song birds have a specialized network of neurons in their brains dedicated to song learning and production (Nottebohm, Stokes, and Leonard 1976). The cell bodies of these neurons are organized into a series of clusters referred to as song control nuclei. The axons of these neurons project to adjacent song control nuclei to form synaptic connections. Lesion studies, electrophysiological recordings, and histological studies reveal that these nuclei form two circuits, an anterior forebrain pathway involved in song learning, and a posterior motor pathway involved in song production (reviewed by Brenowitz and Kroodsma 1996). Nuclei of the anterior forebrain pathway undergo several changes during the period of song learning. This observa tion suggests that learning ceases when neurogenesis is complete (Not tebohm 1981). These observations indicate a different mechanism from the one mentioned above; namely, learning involves simplifying connectivity between neurons (Changeux and Danchin 1976). This model suggests that unused synapses are elim inated during song learning, thus paring down the initial network of con nections. A similar model was proposed for imprinting in chickens (Wallhausser and Scheich 1987). Before we can begin to understand the neural basis of song learning and regulation of the sensitive period, we must distinguish changes in the brain that are a consequence of maturation from those that are func tionally related to learning, regardless of age. The critical question is whether the neural changes described above would be observed if birds were prevented from learning to sing. One method compared the neural development of deafened and hearing birds of the same age (Aamodt, Nordeen, and Nordeen 1995; Burek, Nordeen, and Nordeen 1991). The other compared the brains of tutored birds that had begun practicing song with those of song-deprived birds of the same age that had not begun to sing (Wallhausser-Franke, 69 the Neural Basis of Song Learning in Birds Nisdorf-Bergweiler, and DeVoogd 1995). Both studies uncovered few differences in the brains of control and experimental groups, indicating that most neural changes are likely to be developmental rather than a cause or consequence of song learning. The number of dendritic spines was signi cantly smaller in birds that had learned to sing, supporting the idea proposed by Changeux and Danchin (1976) that learning may involve pruning unused connections between neurons. With the exception of mimics such as starlings and mockingbirds (Mimus polyglottos), most birds, when given a choice, prefer to learn the songs of their own species. Vocal learning was studied in the laboratory with sparrows collected as nestlings before they had an opportunity to learn songs. The birds learned readily from taperecordings during the sensitive period that extends from one to four months of age. Work with sparrows and other species uncovered an interesting paradox: young birds must hear the songs of their own species in order to learn them, but when faced with a potentially confusing array of songs, they are able to select the ones of their own species to serve as learning templates. Young birds raised by their parents in the wild presumably hear a range of sounds, including songs of other species that inhabit the same geographic range. One might assume that they decide which sounds to memorize by observing their parents (most likely their fathers) singing. However, laboratory experiments with tutor tapes in which social cues are not available produce the same outcome. Another possibility is that the size and structure of the vocal tract limits the type of sounds that can be produced, predisposing the bird to learn the correct song. Clearly some anatomical restrictions come into play in the sense that it would be impossible for a hummingbird to produce the call of a crow. However, birds can be induced to learn the songs of other species if they are pre vented from hearing the ones of their own, establishing that anatomy of the vocal tract does not dictate learning preferences (Marler 1991). In conclusion, laboratory studies that remove social cues and provide learn able songs, including those of other species, reveal a learning bias that is guided by instinct. One can imagine circuits in the brain that act as feature detectors and, when stimulated in an appropriate combination, cause the song to be selected as a model for vocal learning. Fledgling sparrows are housed alone in soundproof boxes that are out tted with a speaker and a microphone. Once an hour they hear ten repetitions of a taperecording of normal white-crowned sparrow song, the song of another species (song sparrow, Melospiza melodia or savan nah sparrow, Passerculus sandwichensis), or a white-crowned sparrow song that has been experimentally altered. When edglings hear normal white-crowned sparrow song, they give a series of begging calls that, under natural conditions, help their parents locate them for feeding. Thus, by counting the number of begging calls given in response to our altered white crowned sparrow songs, we can learn whether the edglings perceive them to be acceptable renditions of white-crown song or to be of a foreign species. We used this behavior test to compare responses to normal white crowned sparrow song with responses to isolate white-crowned sparrow song, the simple song produced by white-crowns that have not heard other white-crowns sing. The isolate song consisted of a series of whis tles and lacked the trills and buzzes typical of normal song. The isolate song was as effective as the normal song in eliciting begging calls from edglings. One hypothesis to explain the ef cacy of isolate song is that the whistle, universal to all white-crown dialects, acts as a marker for recognition. We tested this hypothesis using songs arti cially constructed by repeating a single white-crowned sparrow phrase such as a whistle, buzz, or trill, while maintaining normal song duration and tempo. If the whistle acts as the critical ag for identifying the white-crowned song, we predicted that it would elicit as strong a response as the normal song, and the trill or buzz would be comparatively weaker. All of the repeated phrase songs, including buzz and trill songs, proved to be as effective as the normal song in eliciting calls from the edglings. Although avian vocal signals are greatly simpli ed compared with human language, these results par allel ndings of studies of language recognition in human infants. Human infants can recognize the phonemes of all human languages, providing them with the capacity to learn any language. They are even able to dis criminate phonemes not used in their local language, although this sen sitivity disappears once they begin to speak (Eimas, Miller, and Jusczyk 1987; Kuhl 1995). It is possible that, rather than relying on note structure, young birds use tonal or temporal qualities to identify songs of their own species.
Vastarel 20 mg generic
Because nervousness interferes with ne motor control medicine number lookup buy vastarel uk, including voice control, singing in key may reveal self-con dence, status, and extroversion. Virtuosic performance of instrumental music may reveal motor coordination, capacity for automating complex learned behaviors, and having the time to practice (which in turn indicates not having heavy parental responsibilities, and hence sexual availability). Melodic creativ ity may reveal learning ability to master existing musical styles and social intelligence to go beyond them in producing optimally exciting novelty. These indicator functions for music are all speculative, but well established empirical methods are available in biology for testing indi cator hypotheses. Second, one can look for individual-level effects by experimentally manipulating the underlying trait and measuring its effect on the indicator. Third, one can do experiments on mate preferences to see whether people are more sexually attracted by individuals with higher rather than lower indicator values, and whether they attribute higher underlying trait values to those with high indicator values. None of these empirical studies has yet been done, to my knowledge, to analyze human music as a set of sexually selected indicators. Many such studies would have such obvious outcomes that doing them hardly seems nec essary. But even obvious studies such as those showing that healthier peacocks have larger tails (Petrie, Halliday, and Sanders 1991; Petrie 1992) were critical in demonstrating the importance of indicators in other species. Music as a Set of Sexually Selected Aesthetic Displays Whereas indicators reveal useful information, aesthetic displays play on psychological foibles. The basic idea of aesthetic displays is that mate choice works through animal sensation, perception, and cognition, and these psychological processes sometimes have biased sensitivities that other animals can exploit with their courtship displays. For example, a certain species of bird may eat red berries a lot, so evolves eyes with high sensitivity to red and brains that are attracted by the color. This percep tual bias may affect mate choice, predisposing the birds to mate with others who have red rather than blue or yellow plumage. The result would be that the red-biased eyes result in red-biased evolution of courtship plumage (Endler 1992, 1993). Thus many sexually selected aes thetic displays may originate as side effects of perceptual adaptations evolved for other functions. Burley (1988) found that female zebra nches have latent aesthetic pref erences for the red and black plastic leg bands that she used to tag certain males, and not for the yellow or blue bands she put on other males. Of course, male zebra nches of the future will not evolve plastic bands on their legs, but they may very well evolve red coloration if the right muta tions pop up (consider the blue-footed booby of the Galapagos). Thus, almost any perceptual bias that animals have can shape how sexual selection plays out, and which courtship displays evolve in a species. Ryan and Keddy-Hector (1992) and found that these biases are not randomly distributed, but are typically pointed in one direction. With respect to visual traits, for example, all species they inves tigated preferred bright colors over duller colors, larger displays over smaller ones, and higher contrast over lower contrast. With respect to acoustic traits, all species they investigated preferred calls that were louder rather than softer, more frequent rather than less frequent, longer in duration rather than shorter, lower in pitch rather than higher, higher in complexity rather than lower, and with larger repertoire sizes over smaller repertoires. The relevance to sexual selection for music is obvious: any acoustic preferences that our ancestors had could have been exploited, attracted, and entertained by production of the appropriate musical display. Aesthetic traits tend to be hard to distinguish from indicators, because in almost all cases, perceptual biases push sexual selection in the same direction that mate choice for reliable indicators would. Lower-pitched calls, for example, are reliable indicators of body size, because very small animals cannot physically produce very low pitches. Often, traits may function as both aesthetic displays and as indicators (Miller 1997a, 1998; Miller and Todd 1998). The advantage of the aesthetic display theory is that it makes us recognize that any aspect of music that we nd appeal ing could also have been appealing to our ancestors, and if it was, that appeal would have set up sexual selection pressures in favor of musical productions that ful lled those preferences. Fisher considered situations in which both mate preferences and courtship traits are heritable and asked what would happen to both over evolutionary time. He observed that if peahens varied in the length of tail they preferred, and if peacocks varied in their tail lengths, they would end up mating assortatively, with length obsessed females mating most often with the longest-tailed males. Their offspring would tend to inherit genes for both long-tail preference and for long tails at above-average frequencies. If the population had an initial bias, with more females preferring long tails than short, and with more females wanting long tails than long tails were available, this assor tative mating effect would set up a positive-feedback loop between mate preference and courtship trait, leading to ever more extreme preferences and ever more exaggerated traits. Any psychological mechanism used in mate choice is vulnerable to this runaway effect, which not only makes the displays that it favors more extreme, but makes the emotions and cognitions them selves more compelling. If music that moves emotionally or inspires spiritually tended to attract sexually as well over ancestral time, sexual selection can explain its appeal at every level. As a tool for activating certain emotional states, however, music is much better than language. Combining the two in lyrical music such as love songs is best of all as a courtship display. Music has many features that can be interpreted as aesthetic displays that ful ll preexisting perceptual and cognitive preferences. Rhythmic signals are known to be capable of optimally exciting certain kinds of recurrent neural networks as found in mammalian brains. Tonal systems, pitch transitions, and chords probably play on the physical responsive ness of auditory systems to certain frequency relationships. Musical novelty attracts attention by violating expectations, overcoming habitu ation and boredom, and increasing memorability. Music with lyrics reaches deep into cognition through the media of language and imagination. As with indicators, biology has developed empirical methods for demonstrating aesthetic displays that could be extended to human music. The rst step is to perform perceptual experiments to explore the pref erences of receivers for various types of stimuli, charting out which ones are optimally exciting and attractive. For example, vary the beats per minute of a musical stimulus and see which rhythmic speeds best excite various feelings in people. The second step is to measure stimuli actually 344 Geoffrey Miller produced by conspeci cs to see how close they come to being optimally exciting given these preferences. For example, measure the beats per minute in a large sample of commercially produced song and see whether the speeds match the optimal responsiveness curves of human receivers. Many such experiments are pretty obvious, but they become more interesting if they are extended across closely related species to see whether the preference is phylogenetically ancient, or whether it evolved to an extreme form through runaway selection in one species but not in others. For example, if humans respond best to dance music played at 120 beats per minute, but chimpanzees and gorillas do not respond dif ferently to different rhythmic speeds, we would have some evidence for runaway selection affecting rhythmic preferences in the human lineage. Computer simulations of evolution under sexual selection may also prove useful in showing how aesthetic displays evolve. My colleagues Peter Todd and Greg Werner extended our previous sexual selection simulations (Miller and Todd 1995; Todd and Miller 1993, 1997; Todd, this volume) to model the evolution of musical complexity and variety under mate choice (Werner and Todd 1997).
Order vastarel 20mg without a prescription
Thus a lesion in this area results in word blindness (alexia) medications 563 purchase vastarel canada, in which a person can see well but cannot read (and therefore cannot write). Area 39 is located in the pari etal lobe, as is area 40 (supramarginal gyrus) directly in front of it. Both areas send and receive information from many other cortical regions; that is, they are association cortex. Area 40 is crucial for understanding 200 Dean Falk the symbolism of language, so much so that a person with damage here may be mute. Thus, damage in this area results in agraphia, which, as noted, may also be caused by damage to area 39. The cerebellum is the great motor coor dinator that sits beneath the occipital lobes. Its evolutionarily newest part, the neocerebellum, has connections with the frontal lobe and is active during thoughtful speech. This nding is not only recent (Petersen and Fiez 1993), but surprising, because the cerebellum was previously believed to have nothing to do with higher thought. Many other parts of the brain, of course, participate in sophisticated cognition, including various aspects of language. It also should be stressed that, although most language processing takes place in the left hemisphere of most people, the right hemisphere comprehends a certain amount of language and participates in this activity to an extent, for instance, by under standing and providing intonations of utterances (tone of voice, or prosody). Just as the major or dominant left hemisphere is well known for its language abilities, the minor right hemisphere is traditionally regarded as the musical part of the brain. Because recent advances in medical imaging technology make it possible to investigate the brains of subjects as they engage in various musical activities, the relationship between 201 Hominid Brain Evolution and the Origins of Music language and music is open to new exploration and, potentially, new surprises. Neuroanatomical Substrates of Music Although all normal people are competent in at least one language, not everybody is a pro cient musician. Investigations pertaining to neuro logical processing of music are therefore complicated by the fact that some focus on musically trained subjects whereas others deliberately select subjects that are musically naive. Despite the fact that differences between the groups can be illuminating, studies of musi cians are particularly relevant for questions pertaining to the speci c components of perceiving and producing music. Listening to , reading, or playing the partita each recruited speci c cortical areas. Each of the ten musicians initially listened to and then played ascend ing and descending scales on the keyboard with their right hands. Merely listening to scales activated area 42 in both hemispheres and area 22 on the left (indicated by stars in gure 13. When subjects played the scales themselves, the right cerebellum (star) that connects with the left frontal lobe became activated. As noted, the neo cerebellum (especially on the right) is also engaged during thoughtful speech. In addition, playing scales stimulated portions of the left premotor cortex (stars in area 6) that, again, appear to overlap with language areas; that is, the left pre motor cortex involved in writing with the right hand (Xs). Clearly, the perception and manual production of simple scales share some neuro logical substrates with the perception and manual production of simple words (Sergent et al. When a musical score is simply read without listening or playing, the acti vated area of the brain is not 39 on the left as is the case when words are read, but rather part of visual area 19 (star) on the left (in addition to visual area 18 bilaterally), which is important for spatial processing, interpreting where rather than what a visual stimulus is. This makes sense because pianists read notes not as isolated items but in terms of their 202 Dean Falk positions relative to one another (Sergent 1993). Listening to a score, as opposed to hearing scales, adds area 22 on the right side, af rming that listening to music involves differential activation of the right hemisphere, as is widely believed. Of interest, reading a score while listening to it adds another area to those that are stimulated by each activity alone, namely, the top part of the supramarginal gyrus (area 40) in both hemispheres (star and cross-hatching) that, on the left, is profoundly important for grasping the symbolism of language. In addition to the areas outlined above, two areas are recruited that are not activated by any of these activities alone. One of these, the superior parietal lobule, or area 7 (star), is acti vated in both hemispheres. For example, a blindfolded pianist with a lesion in part of area 7 on the left would not be able to identify a piano key as such by touching it with his right hand, although he would know that it was cool, relatively small, and moved when pressed, but these sensations would not be synthesized into the concept of a piano key. In addition to the left motor and premotor and right cerebellar acti vation that one expects from the simple playing experiment described above, another frontal lobe area is recruited in the left hemisphere in the reading-playing-listening task. This nding makes perfect sense from a neurological perspective when one considers the organization of the primary motor cortex. As such, it would be interesting to learn if this region is speci cally involved in production of American sign language that, like lan guage, is left-hemisphere dominant and dissociated from nonlinguistic gesture (Hickok, Bellugi, and Klima 1996). First, they document the existence of a dis tributed neural network that incorporates specialized nodal regions for processing sensory and motor aspects of music, as is the case for lan guage. Second, in some cases, cortical areas that underlie musical activi ties have been shown to be next to , and partly overlap with, those engaged in similar language tasks. Finally, portions of certain areas are differentially activated in the left hemisphere during speci c musical activities; for example, area 22 from listening to simple scales, part of area 19 with simple reading of a musical score, motor and premotor (area 6) cortices (plus right cerebellum) during simple playing of the keyboard with the right hand, and the top part of area 44 during the multifaceted sight-reading task. Thus, musicians, at least, rely a good deal on their left hemisphere when processing music. Again, area 42 became activated bilaterally and area 22 was more responsive on the right. Deeper brain structures that contribute to the auditory ascending pathway were also activated (medial geniculate nuclei, inferior colliculus, lateral lemniscus), causing the authors to conclude that imagining music and actually hearing it activate the same neurological substrates. They also noted that another deep structure, the putamen, which is activated on the left, may be involved with timing of the imagined music. This study is important because it is one of the few that imaged deep brain structures during musical cognition. If there is a surprise in the above recent ndings, it is the extent to which musical activities engage the left hemisphere in a manner that par allels the processing of language. For one thing, as noted, the right temporal association cortex (area 22) is recruited on rst hearing a piece of music (Sergent et al. It thus appears that melody has emotional content for both language and music; recall that limbic structures that process emo tions were activated on the right as volunteers imagined music (Chen et al. Prefrontal cortex is generally known to be important for keeping information in mind during goal-oriented tasks. In general, the right hemisphere is also more sensitive to harmony (Tramo and Bharucha 1991), concordant with its proclivity for recognizing and producing har monic ratios within complex tones (Preisler, Gallasch, and Schulter 1989). Music Meets Speech: Singing in the Brain Given the widely held view that the left hemisphere is dominant for lan guage and the right is superior for music, it seems obvious to ask about the neurological substrates for the one activity that clearly incorporates both endeavors, namely, singing with words. In what may be the rst experiment to investigate hemi spheric dominance for singing both with and without words, Cadalbert et al. Singing with words is associated with wider right-side lip opening than is the case for singing without words, indicating involve 205 Hominid Brain Evolution and the Origins of Music Fig.
Discount vastarel 20mg overnight delivery
Meeting such obvious similarities treatment 32 for bad breath purchase vastarel 20 mg online, an ethnomusicologist will often try to discover along which tracks they must have been circulating and trace them back to one common source. In my opinion it is quite unlikely that any relationship can be proved during historical times between Taiwan and Niger, or between Albania and Sulawesi. If we imagine that such likeness may refer not to historic relationships but to the supposedly common origins of humans, it seems that the two types of explanations differ little (through diffusion or through spontaneous similarities) between cultural history or natural innate schemes. Because if such close similarities, in music just as in mythology, are the only surviving tokens of an ancient diffusion, the question is, why have only these features seemed to survive On the other hand, if they are not the result of forgotten migrations but of a natural scheme, problems related to geography and history no longer exist, and thousands of years count for nothing in evolutionary terms. The main problem is to under stand how precise sound organizations can be inscribed in every brain, and how musical choices emerge from them or deal with them. I leave it to psychologists and neurophysiologists to explain the muscular and neural laws that help us understand the ubiquity of certain tempos and rhythms in animal vocalizations and human music. To support my hypothesis of universals given by nature in music, I will illustrate several similarities between animal and human signals (see Mache, F. Culturalists claim that one may not apply 476 Francois-Bernard Mache the same categories to different cultures, and even that the de nition of what is the same is always a matter of cultural relativity. In the same manner, some psychologists claim that it would be anthropocentric, and therefore wrong, to assimilate or even to compare animal and human sound features. By using this term, one refers to likenesses that, strong as they may seem, have no explanatory value, because they refer to separate causal series. The question is, can acoustic features that are common to animals and humans be viewed as simple conver gences, with no scienti c value because they contradict many other dif ferences The question of universals in music is directly related to the question of its origins. Being a musician rather than a biologist, I tend to observe surface structures, musical features. If they correspond to concepts provided by evolutionary theories, one can state that the universal and the biological coincide. To propose an answer, I submit a number of examples taken from several animals, illustrating categories that are considered typically musical. I borrow both terms from biologists, with a slightly different interpreta tion if musical genotypes should turn out to be less constraining than their counterparts in biology. In my career as a composer I was interested in phenotypes long before I undertook to connect them with possible genotypes. It is only after long acquaintance with animal models, which I have used in many works since the beginning of my career in 1958, that I wondered why I could so will ingly perceive some latent music in the sounds made by whales, frogs, crickets, and birds. Eventually I perceived the correctness of the mythic tradition that presents music as related to bird song. What is new about this antique intuition is the taperecorder, which allows us to compare and to verify. The objection that bird song is only the expression of biological functions, like territorial defense or courting, and belongs to the semi otic sphere, not to the aesthetic, is not as weighty as it seems. I mention it now to indicate that it did not prevent me from looking for a natural justi cation for my use of natural models. An important family of rhythms among the different musical systems is the aksak, which exist in a very large area corresponding to the empire of Alexander the Great, from the Balkans to the Pamirs. They oppose an 477 Necessity of and Problems with a Universal Musicology irregular number of basic units, very often grouped by three and by two. Examples are seen in the songs of Tockus erythrorhynchus, the red-billed hornbill, and Alectoris rufa, the red-legged partridge. For example, in this song by Turtur brehmeri, a blue headed dove, the rst two notes of the accelerando are separated by 2. What is most universally considered as musical is the occurrence of a set of discrete pitches. Many mythic traditions, in Greece and China, for example, attribute this essential cre ation to a god or a cultural hero. Halcyon badius, the chocolate backed king sher, moves up and down along his own scale, character ized by very small intervals. More subtly, Cossypha cyanocampter, the blue-shouldered robin-chat, is not satis ed with enumerating the tones of its scale, but operates on it by building melodic motives as elaborate as many human achievements, and even sounding so close to them that one might be mistaken. The same melodic use of a scale, but in this case a kind of chromatic scale, occurs in Erythropygia leucosticta, the north ern bearded illadopsis. In the example of Trichastoma albipectus, the scaly breasted illadopsis from Kenya, you get a legato instead of a previous staccato. Still closer to human organization is the evidence for a hierarchy between the degrees of a scale. A note may assume a particular role according to its frequency and position in the melody. This is true for human systems, such as the tonic and dominant in tonal systems, or the shahed or forud in Iranian dastgah. In the songs of Erythropygia leucophrys, the white-browed scrub robin, a kind of keynote appears at the end of each stanza. The process of transposition is of particular relevance for a compari son between animals and humans. It implies memory for and conscious ness of a given sound pattern treated as a whole. Whenever a sound model is imitated by a bird whose range does not t, it is transposed both 478 Francois-Bernard Mache in pitch and duration, as for example when Lanius minor, the lesser grey shrike, imitates a rooster. Clearly, in many cases the syntax of animal signals has something in common with music. My view that we are dealing with a functional similarity in animal species and human often meets some objections, which can be summa rized this way: animal sound signals belong to pure semiotics. My answer is rst that the idea of a gratuitous aesthetic pleasure is but a very small part of musical behavior in humans. It took on special impor tance only one or two centuries ago, in European civilization. It is quite a naive idea to consider music only as the thing a young lady does when per forming a piece on her piano, with friends and family attending. The Toradjas of Sulawesi never make music for the sole pleasure of singing or listening; they have no lullabies, no wedding songs, no dirges. It would be bold to say that they have no music simply because this activity gures in social occasions where singing is just part of the whole. It is not proved that such singing neighbors avoid territorial con icts more easily than those that sing alone or ignore each other. I would rather suggest that the opinion maintained by several biolo gists such as Thorpe (1966) is right: there is also something like an intrin sic pleasure in singing. The luxurious display of some of the best singers suggests that they go far beyond the signals that would be necessary for keeping a territory or mating. The views that the ethologist Sebeok (1975) expressed seem to support such a hypothesis, which I submit to more expert specialists. It implies that the whole elaboration of a culture, meaning a collective structure of symbolic imagination, might stem from this lavishness of nature exceeding its limited basic pur poses.
Discount 20 mg vastarel with visa
This is a tricyclic antidepressant but also has significant anticholinergic effects symptoms hiv buy cheapest vastarel. In the differential diagnosis of dementia is the syndrome of pseu dodementia caused by depressive syndromes. Some of the characteristics of depressive pseudodementia include manifestations of depressive syndromes such as withdrawal from social behaviors, disruption of the sleep wake cycle, and altered eating behavior. A good behavioral clue to pseudodementia versus true dementia is the type of answers given on mental status testing. Paraphasic errors: the production of unintended syllables, words, or phrases during the effort to speak. Nucleus basalis of Meynert: A group of nerve cells that has wide projec tions to the neocortex and is rich in acetylcholine and choline acetyltransferase. Multi-infarct dementia: Dementia in the setting of cerebrovascular dis ease, occurring after multiple cerebral infarctions, whether large or small (lacunar). When the anterior half of the cortex is affected, it tends to produce problems with behavior, executive dysfunction. Of tremendous practical importance is the fact that virtually all dementias that are treatable for cure or at least may be amenable to slowed progression are subcortical dementias. The amyloid plaques stain positively with antibodies to amyloid precursor protein. There are known mutations in the amyloid precursor protein as well as two homologous pro teins, presenilin-1 and presenilin-2 that tend to present with early-onset disease. As a correlate, functional imag ing studies show hypometabolism in the temporal and parietal cortices (Fig. There seems to be particular degeneration of the cholinergic cells that project to the cortex from the basal forebrain, particularly the nucleus basalis of Meynert. One of the conse quences of cholinergic loss is also extreme sensitivity to the deleterious effects of anticholinergic medications. Differential Diagnosis If cognitive decline occurs with prominent mood disturbance, then one con sideration is depression or pseudodementia. It is often difficult to distinguish which occurred first, because many elderly patients with cognitive decline and declining level of independent functioning suffer from a reactive depression. If the patient has a history of irregular stepwise decline in functioning, especially if the patient has had apparent stroke symptoms or transient ischemic events, or has known cardiovascular disease or atrial fibrillation, then multi-infarct dementia is the most likely diagnosis. Axial T1-weighted magnetic resonance images of Alzheimer disease patient showing bilateral hippocampal atrophy and generalized atro phy. Other common causes of dementia include cognitive decline caused by long-standing alcoholism, or dementia associated with parkinsonism. Both of these underlying conditions are readily discovered by the appropriate asso ciated medical history. The classic triad is dementia, gait disturbance, and urinary or bowel incontinence. Relief of hydrocephalus through placement of a ventriculoperitoneal shunt can reverse the cognitive decline. Donepezil (Aricept) and rivastigmine (Exelon) are cholinesterase inhibitors that are effective in improving cognitive function and global clinical state. Other issues include wakefulness, nightwalking and wandering, aggression, incon tinence, and depression. The Alzheimer Association is a national organization developed to give support to family members and can be contacted through In its early stages, affected patients have a normal neurologic exam except for the mental status exam ination and olfactory testing. The association between delirium and cog nitive decline: a review of the empirical literature. A neuropsychologic examination revealed changes in exec utive functioning (verbal fluency, attention, judgment) and difficulties performing previously learned tasks (dyspraxia). Based on the temporal progression of dementia and behavioral decline, then sleep-related disorders and parkinsonism, diffuse Lewy body disease is the likely diagnosis. Her history is also sig nificant for clinical worsening with an empiric trial of a psychotropic, not oth erwise specified. With Parkinson disease, patients can also have a dementia, but it is predomi nantly subcortical (slowed thought processes, retrieval, attention, and concen tration) and is usually not a predominant clinical feature of the disease. Often the dementia and motor dysfunction are more global and include multiple cortical and subcortical deficits, extrapyramidal and pyramidal dysfunction, and a more rapid clinical course. Executive function: Mental capacity to control and plan mental skills, the ability to sustain or flexibly direct attention, the inhibition of inappro priate behavioral or emotional responses, the planning of strategies for future behavior, the initiation and execution of these strategies, and the ability to flexibly switch among problem-solving strategies. Ideomotor apraxia: Disturbance of voluntary movement in which a per son cannot translate an idea into movement. Whether this difference is clinically useful is under investigation, as is the diagnostic utility of functional imaging. Cognitive and behavioral symptoms have been shown to improve with this class of medications, however, not depression. Haloperidol is a dopamine-receptor blocking agent that can have severely deleterious consequences in this disorder. Practice parameter: evaluation and treatment of depression, psychosis, and dementia in Parkinson disease (an evidence-based review): report of the Quality Standards Subcommittee of the American Academy of Neurology. There was slight limitation of head movement to either side, but no pain with neck extension. He was able to stand with his feet together with his eyes open, but he nearly fell when his eyes were closed. Understand the range of pathologic and clinical manifestations of vitamin B12 deficiency. Considerations the pertinent features of this case include the presentation, unsteadiness of gait, and numbness and stiffness. There was a stocking decrease in sensation, specifically vibration and joint position sense, which strongly suggests a neuropathy involving myeli nated fibers. Involvement of the dorsal columns of the spinal cord, at or above the lumbar level is also a possibility. The ataxic heel knee to shin maneuver, also points to aberrant input to the cerebellum, which comes through large fibers. All of these findings suggest involvement at multiple levels of the nervous system. The imaging study confirms involvement of myelinated regions in the spinal cord, specifically the dorsal columns and in the brain. Another clue on physical diagnosis is the abnormal tongue and prematurely graying hair. The spinal cord is a tubular structure originating from the medulla of the brain and extending through the bony spine to the coccyx. Ascending sensory and descending motor white matter tracts are located peripherally; posterior columns govern joint position, vibration and pressure, lateral spinothalamic tracts pain and temperature, and ventral corticospinal tracts carry motor signals. There is a diffuse effect on the spinal cord, prima rily the posterior lateral columns, explaining the early loss of vibratory sense. Cyanocobalamin is a compound that is metabolized to a vitamin in the B complex commonly known as vitamin B12.